



E – 75 800 à 72 000 AEC2
Primiglaciaire – interstade d’Odderade
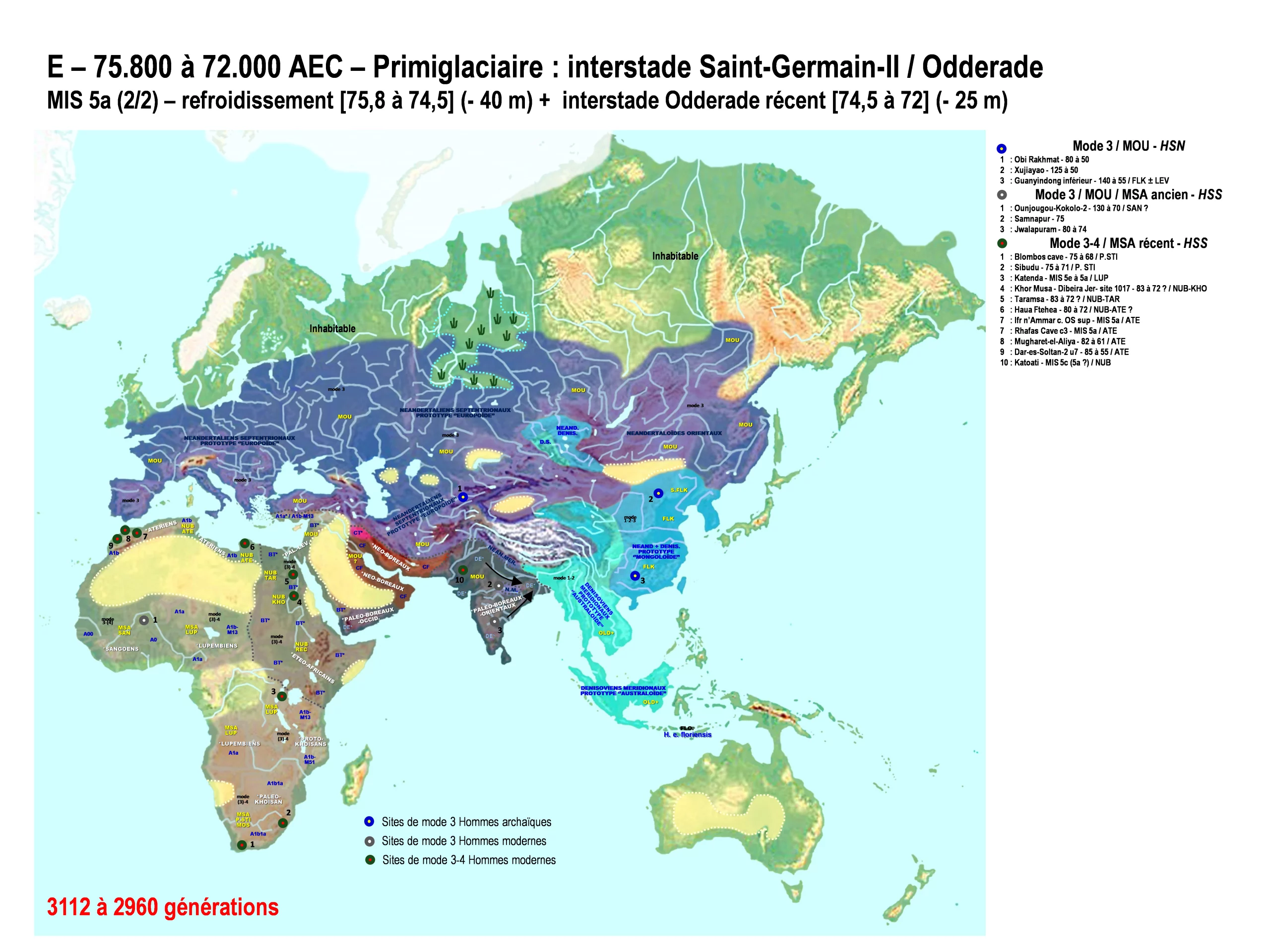
Glacial intra-Odderade : 75.800 à 74.500 AEC
Odderade récent : 74.500 à 72.000 AEC
MIS 5a (2/2)
E – 75.800 à 72.000 AEC – Primiglaciaire : interstade Saint-Germain-II / Odderade
Climat
La partie ancienne de l’interstade d’Odderade se termina par un refroidissement marqué qui apparait beaucoup plus clairement dans les carottes glaciaires du Groenland (GS-21) que dans les séries sédimentaires continentales. Cette période froide au sein d’un interstade pourrait être appelée glacial intra-Odderade ; elle dura de v. 75.800 à 74.500 AEC. Elle fut suivie par un nouveau réchauffement que nous appellerons interstade d’Odderade récent (GI-20) et qui dura environ 1300 ans, de v. 74.500 à 72.000 AEC. L’Odderade récent fut la dernière période du MIS 5. Le niveau moyen des mers oscilla peut-être aux alentours de – 40 mètres au cours du glacial intra-Odderade et de – 25 mètres au cours de l’Odderade récent ?
Au cours des périodes clémentes de l’Odderade, l’humidité fut moins importante que lors du Brorup, mais demeura suffisante pour maintenir des paysages arborés au Nord de l’Europe. Le Sud de l’Europe fut moins affecté par le froid que les régions situées au Nord des Alpes, ce qui permit le maintien des faunes locales.
En Afrique, la phase humide Pré-Maluekienne se poursuivit.
Peuples d’haplogroupes A et BT
C’est peut-être dans cette seconde partie de l’Odderade que les industries nilotiques Taramsienne (NUB TAR) et Khormusienne (NUB KHO) se différencièrent vraiment, peut-être portées par des individus BT* ? Des similarités ont été pointées entre le Taramsien et les industries MSA du Sud de l’Afrique, peut-être simplement parce qu’elles contenaient les unes et les autres des marqueurs industriels MSA standards ? Les dates de ces industries sont toutefois difficiles à préciser, même si les dates anciennes que nous avançons semblent aujourd’hui les plus crédibles. En suivant cette logique et en prenant acte que le Nil s’arrêta vraisemblablement de couler en continu pendant toute la durée des MIS 4 et MIS 3, nous proposons de ne pas prolonger ces industries au-delà de 72.000 AEC et donc de les faire disparaître à la fin du MIS 5a.
Peuples d’haplogroupes DE
- Les populations *Paléo-Boréales-Occidentales, porteuses de l’haplogroupe DE*, occupaient vraisemblablement l’Arabie méridionale au cours de l’Odderade. Ce n’est que lorsque le niveau des océans baissera – au début du MIS 4 – que certains de leurs groupes rétromigreront en Afrique où l’haplogroupe E émergera plus tard [cf. carte F].
- Favorisées par le climat relativement clément de l’interstade d’Odderade récent et par le fait qu’un Monde vide d’Hommes modernes leur épargnait une compétition marquée, les populations *Paléo-Boréales-Orientalescontinuèrent vraisemblablement d’étendre l’haplogroupe DE* le long des côtes méridionales de l’Inde ainsi qu’à l’intérieur du sous-continent. Après l’Inde – où les derniers Néandertaloïdes locaux subsistaient certainement encore dans des zones refuges – ils parvinrent peut-être dans la zone des Sundarbans au seuil de l’Indochine ? Aujourd’hui, les intouchables Adivasi du Bengale Oriental expriment toujours cet haplogroupe DE* qui est rare par ailleurs.
Peuples d’haplogroupes CF
Frères des DE, les porteurs de l’haplogroupe CF – et locuteurs des langues *Néo-Boréales [cf. carte C & D] – étaient probablement localisés au Nord-Est de la péninsule arabique et au Sud de l’Iran, près des rivages du Golfe Persique, dans ce que nous avons appelé le ‘’hub’’ moyen-oriental. Cette région – qui avait constitué le débouché naturel des premiers Humains modernes sortis d’Afrique via l’Arabie – était particulièrement intéressante parce qu’elle demeurait perpétuellement accueillante, même au cœur des épisodes glaciaires les plus froids et les plus secs. Etant donné le devenir des haplogroupes qui sortiront bientôt de lui, il est difficile d’assigner une autre position géographique au groupe CF qui, faute de vestiges, reste encore pour nous un groupe théorique mais dont l’existence fut génétiquement incontestable. Au cœur de cette ‘’tête de pont’’ eurasienne, les Hommes modernes CF densifièrent leur population et commencèrent à diversifier leurs haplogroupes ADN-Y. Il faut donc comprendre qu’à l’instar de l’Afrique du Sud où les Khoisans actuels concentrent l’essentiel de la variabilité humaine mondiale actuelle, les premiers Humains modernes d’Eurasie concentraient, en germe, l’essentiel de la variabilité humaine non africaine dans la région du Golfe Persique. A partir du cœur de ce ‘’hub’’ moyen-oriental, des pulsations migratoires emporteront bientôt au loin les porteurs de certains variants haplogroupaux de CF qui emmèneront leurs langues avec eux ; cela dans les trois seules directions possibles : la voie du Sud(Est) vers l’Inde et l’Extrême-Orient (puis de là, en Océanie ou en Amérique) ; la voie de l’Ouest vers le Proche-Orient et l’Anatolie (puis, de là, en Afrique du Nord ou en Europe) ; et – lors des périodes suffisamment clémentes – la voie du Nord vers l’Asie centrale (puis, de là, dans les steppes asiatiques et en Amérique). La quatrième direction théoriquement possible – la rétromigration vers l’Afrique – sera empruntée par des tribus DE* au début du premier maximum glaciaire, puis se fermera derrière eux en raison de la désertification qui affecta la péninsule arabe pendant tout le MIS 4 [cf. carte F à H].
Peuples Paléo-Levantins
Bien que nous ne disposions pas de vestiges humains levantins datés de l’Odderade, la persistance d’une population moderne *Paléo-Levantine est plausible, avec A1a*, A1b-M13 et BT* comme haplogroupes majoritaires putatifs. Rien ne permet d’affirmer que ces populations modernes du Proche-Orient étaient restées cantonnées au seul Levant et à la côte de Cyrénaïque ? Peut-être certains groupes s’étaient-ils installés sur la côte méridionale de l’Anatolie ?
Hommes archaïques
Aux époques que nous étudions, le Monde eurasien était certes vide d’Hommes modernes, mais il n’était pas vide d’Hommes archaïques. Dans ce cas, pourquoi ces derniers n’ont-ils pas légués leurs propres haplogroupes ADN-Y à leurs descendants métissés ? La réponse découle de leurs fonctions cognitives moins développées dont résultaient plusieurs conséquences néfastes : 1) elles interdisaient une forte densité démographique dans une Nature non maitrisée où l’Homme demeurait à la fois un prédateur et presqu’une proie comme les autres ; 2) elles entrainaient une moindre capacité de résistance à la pression colonisatrice des envahisseurs modernes ; 3) elles offraient une chance quasi inexistante aux hommes (mâles) archaïques d’avoir des enfants avec des femmes modernes ! En effet, même avec des cognitions pleinement modernes de part et d’autre, l’histoire récente des colonisations a montré qu’il est exceptionnel que des hommes indigènes – techniquement sous-développés – fassent des enfants avec des femmes immigrantes et donc exceptionnels qu’ils lèguent leurs haplogroupes ADN-Y ; et si cela se produit quand même, il est exceptionnel que les fils nés de telles unions se retrouvent en position d’engendrer de nombreux descendants parce qu’il est exceptionnel qu’ils accèdent aux responsabilités depuis leur position d’hommes stigmatisés. Ce sont toutes ces contraintes qui ont rendu presque impossible la pérennisation sur le long cours des haplogroupes ADN-Y indigènes. En revanche, dans ces mêmes situations de colonisation, il est très fréquent que des hommes migrants fassent des enfants à des femmes indigènes ; naturellement, même s’ils sont socialement stigmatisés eux-aussi, les garçons métis nés de ces unions portent l’haplogroupe ADN-Y de leur père migrant et leurs éventuels descendants continueront à véhiculer cet haplogroupe sur le long terme. Quelle que soit la justesse de ce raisonnement, force est de constater qu’on n’a encore jamais trouvé un haplogroupe ADN-Y archaïque dans le Monde d’aujourd’hui.
Peuples, langues et halpogroupes ADN-Y
Revenons sur un point déjà abordé précédemment, parce qu’il est important pour comprendre le peuplement du Monde. On pourrait légitimement s’étonner de l’association étroite qui est faite dans cet atlas entre un groupe ethnolinguistique préhistorique et un haplogroupe ADN-Y qui lui est précisément rattaché. On croyait pourtant savoir depuis longtemps qu’il était parfaitement vain de chercher à rattacher des langues à des gènes, et plus particulièrement encore à des haplogroupes ADN-mt ou ADN-Y ; cela parce qu’il est facile d’observer que les groupes ethnolinguistiques actuels sont toujours composés de femmes et d’hommes porteurs de plusieurs haplogroupes différents, même s’il arrive souvent que l’un de ces haplogroupe prédomine sur tous les autres !
Mais cette observation ne suffit pas pour mettre un terme au débat. En effet, nos groupes ethnolinguistiques actuels sont le résultat de mouvements protohistoriques et historiques qui ont eu pour résultante d’amalgamer des populations d’origines diverses, dans une succession de melting-pots portés à l’incandescence par la guerre, la coercition et l’asservissement [cf. Introduction de l’atlas n°3 & atlas n°4 où cette question est détaillée].
Or, la situation était bien différente au début de la colonisation du Monde, lorsque les premiers Hommes modernes se répandaient hors d’Afrique dans des régions encore vides d’autres Hommes modernes ; où ils ne rencontraient que des Hommes archaïques – non-locuteurs ou faiblement locuteurs – qui s’effaçaient rapidement devant eux. Dans la mesure où les groupes de chasseurs-cueilleurs étaient de très petite taille et de structure clanique patrilocale, les haplogroupes ADN-Y des chefs polygynes devenaient rapidement ceux de tout leur clan (ex : installation hégémonique de CT au seuil de l’Eurasie). Ainsi, quand il arrivait qu’un jeune chef (i.e. en capacité d’avoir de nombreux fils) porte – par mutation – un nouveau variant ADN‑Y, ce variant se répandait rapidement dans toute sa petite tribu ; puis, en cas de victoire sur les tribus voisines, ce variant marginalisait tous les autres variants parallèles (ex : DE ou CF qui marginalisèrent les autres variants de CT, que l’on appelle collectivement CT* pour signifier qu’ils n’étaient ni-DE ni-CF). Ensuite, pour peu que les descendants patrilinéaires de ce chef ‘’mutant’’ continuent d’accumuler une série de succès guerriers, son haplogroupe fondateur se répandait dans toute une région et pouvait même y devenir rapidement la norme [cf. introduction]. Bien sûr, puisque de nouveaux variants ADN-Y se produisent sans cesse, des tribus filles pouvaient tôt ou tard reproduire le même processus à partir d’un nouveau chef mutant … pour peu qu’elles aient la chance de surfer elles aussi sur le succès guerrier et donc reproducteur (ex : DE et CF éclateront bientôt en de nouveaux variants qui reproduiront fractalement la même histoire). Dans ce processus, le départ d’une tribu dans une nouvelle région dépourvue d’Hommes modernes conduisait inévitablement cette nouvelle région à être peuplée par un seul et même haplogroupe ADN-Y (effet fondateur). Tous les haplogroupes du Monde actuel – et tous les haplogroupes anciens que nous reconstituons génétiquement pour les relier – ont été des variants apparus par mutation chez un unique garçon bien réel dont le succès reproductif à long terme fut absolument gigantesque : soit parce que lui-même ou ses très proches descendants éliminèrent leurs concurrents masculins dans leur région d’origine, soit parce qu’ils partirent fonder une nouvelle nation au loin. Il faut comprendre que bien d’autres variants différents sont aussi apparus chez de très nombreux autres garçons ; mais ceux-ci n’eurent pas le même destin reproducteur que leur cousins fabuleux, et ne purent jamais donner naissance à de grandes lignées ADN-Y ; pour la bonne raison qu’eux où leurs très proches descendants mâles ne furent jamais de grands chefs victorieux dont les fils et les petits-fils connurent à leur tour la victoire, et qu’ils ne furent pas, non plus, de grands explorateurs de l’inconnu. Audaces fortuna juvat !
Plus tard, lorsque des chocs frontaux se produiront entre des groupes devenus très divergents, dans un Monde qui commencera à devenir plein, l’association entre un haplogroupe ADN-Y et un groupe ethnolinguistique continuera cependant encore d’exister : parce que la mécanique impitoyable des guerres primitives entraînait le plus souvent le génocide complet des hommes vaincus qui étaient cannibalisés ; parce que le concept d’esclaves masculins était sans intérêt pour des peuples paléolithiques, encore non accumulateurs et non productivistes. La mécanique populationnelle et sociétale qui s’enclenchait dès lors, a été longuement détaillée dans l’introduction.





